
概要
临床特点.
Y染色体不育的特点是无精子症(无精子),重度少精子症(<1x106精子/ml精液),中度少精子症(1-5x106精子/ml精液),或轻度少精子症(5-20x106精子/ml精液)。Y染色体不育的男性虽然体检可发现有小睾丸,但通常没有明显的症状。
诊断/检测.
无精子症或少精子症和/或异常精子形态/运动的健康男性,在排除其他导致不孕的原因后,可诊断怀疑为Y 染色体不育症。染色体微阵列(CMA)或常规细胞遗传学检测显示这些男性中的5%-10%有染色体异常。分子检测揭示这些男性中另有5%-13%的人存在Y染色体长臂微缺失。
管理.
对症治疗: 可以通过ICSI(卵胞浆内单精子注射)体外受精而怀孕,在体外受精过程中,精子从射精(男性少精症)或睾丸活检获取(男性无精子症),显微注射到其伴侣的卵子内。
其他: AZFb和AZFa缺失的男性,睾丸取精体外受精是无效的,但是对大多数AZFc缺失的男性来说,睾丸取精体外受精可获得成功;在可体外获取精子的男性中,Y 染色体长臂的存在或缺失对受精或妊娠率无明显影响;出生缺陷风险与其他采用辅助生殖技术助孕的夫妇是相似的。
遗传咨询.
Y染色体不育以Y连锁方式遗传。由于Y染色体微缺失的男性是不育的,因此缺失通常是 新生突变,先证者的父亲是不存在异常的。尽管他们有严重的精子发生障碍,某些AZF区域 缺失的 男性偶有自发生育,但他们的儿子是不育的。这将发生在约4%的重度少精子症的夫妇中,特别是在女性伴侣年轻且生育能力极佳的情况下。经ICSI获得的妊娠中,男性后代会有与其父相同的缺失,具有很高的不育风险。 注意,某些Y缺失,包括最常见的Y缺失(gr/gr),并不一定导致不育,只是不育的一个危险因素。Y染色体缺失父亲的女性胎儿没有 先天性异常或不孕的风险。通过辅助生殖技术(ART)怀孕并已知有导致Y染色体缺失男性的风险,可以进行特定的产前检查或胚胎植入前检查,以确定胎儿的性别和/或Y染色体是否缺失。
诊断
临床诊断
Y染色体不育男性的体格检查通常是正常的,尽管有些会有小睾丸的表现。
应排除其他可导致男性不育的可能(见 鉴别诊断)。
注意,有关Y染色体缺失的临床文献可能非常令人困惑,导致对缺失表型效应的误解。对这个问题阐述最合理的文献是2011年的一篇综述[ Silber 2011 ]。
精液分析. 射精检查以确定精子的数量、活力和形态。精液分析应遵循WHO的指导方针,即检验和处理人类精液的实验室手册[ WHO 2010]。确定了以下精子计数的种类 (Table 1):
Table 1.
精子计数的分类
| 精子计数的分类 1 | 百万精子/mL |
|---|---|
| 无精子症 | 0 |
| 重度少精子症 | <1 |
| 中度少精子症 | 1-5 |
| 轻度少精子症 | 5-20 |
| 正常 | >20 |
在每个类别中,精子的形态和/或运动可以正常或异常 (asthenoteratozoospermia).
检测
染色体微阵列 (CMA). CMV检测是进行外周血染色体不平衡检测,如果有可覆盖异常区域的有效探针,可以检测 非整倍体(如47,XXY)以及Y染色体的缺失或重复。CMA这一分析补充和证实了 分子遗传学检测。值得注意的是,对于Y染色体缺失的CMA数据的解释有时会变得复杂,因为许多与Y不育有关的基因存在多个拷贝且有相似序列。虽然CMA不能检测出平衡的染色体重排,那些与不育相关的拷贝数变异是可以被检测到的,例如 SHOX基因的缺失,这对医疗管理是很重要的[ Jorgez et al 2011]。
细胞遗传学分析. 大约5% - 10%与无精子症/少精子症和或精子形态/运动异常有关的原因不明的不育男性,有 染色体异常发生,通常是性染色体的(即涉及性染色体)异常,也有 常染色体的异常。异常可以是数量上的(如Klinefelter综合征47,XXY)或者是结构上的 (如常染色体平衡 易位)。常规的细胞遗传学研究,包括G显带和荧光原位杂交( FISH)分析,使用Y连锁基因特异性探针对外周血进行分析可以检测以下的Y染色体结构异常,当存在:
然而,单独的细胞遗传学分析不能:
- 确定是否有一个细胞遗传学上可见的延续至AZF区域的缺失。
睾丸活检. 睾丸活检可揭示下列任何一种情况:
- 唯支持细胞综合征 (SCOS),无精子症与生精小管中缺失或仅有的生殖细胞有关,因为大部分细胞都是支持细胞,没有或少有生精细胞
- 精母细胞成熟停滞,但是无精子细胞或成熟的精子
分子遗传学检测
- AZFa区域中一个罕见的USP9Y单基因异常
临床检测
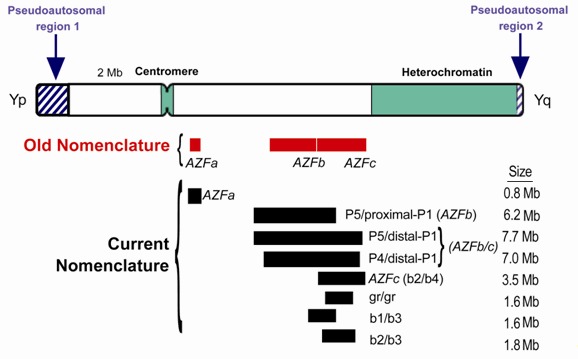
- 缺失/重复分析. Y染色体缺失的分子诊断包括Y染色体内相对广泛区域内的一系列PCR(多聚酶连反应)。根据断裂点的不同,Yq臂间微缺失可导致长臂各部分的丢失。定位于Y染色体的特定基因 (例如, USP9Y, DDX3Y, BPY/VCY, HSFY1, HSFY2, KDM5D, RPS4Y2, RBMY, PRY, DAZ, and CDY; 见 分子遗传学)应该包含在分析中,以评估Y染色体的完整性。缺失/重复分析(如, 定量PCR)可帮助确定基因拷贝数[ Noordam et al 2011]。注意: (1) 尽管检测指南(包括使用阳性和阴性对照样品进行PCR分析)已发布[ Simoni et al 2004],推荐的检测仍是不够的,尤其是与特定基因有关的应包含在内。 (2) 已有设计好的商业化的试剂盒,但需要进一步的评估[Aknin-Seifer et al 2005]。最初,确定了三个AZF区域:AZFa, AZFb和AZFc (无精子症因子a, b和c),从着丝粒到端粒顺序依次排列于Y染色体长臂(Yq),互相没有重叠。然而,后续的研究表明AZFb和AZFc存在重叠[ Repping et al 2002] (见 Figure 1 和 分子遗传学)。因为缺失往往发生在大的回文序列之间,需要一个更适合的关于复发性缺失类型的术语来命名侧翼重复[ Yen 2001 , Repping et al 2002]。
Table 2.
Y染色体不育的分子遗传检测综述
| 基因 1 | 检测方法 | 遗传机制 2 | 表型 | 变异检出率 3 |
|---|---|---|---|---|
| N/A | 缺失/重复 分析 4 | AZFa臂间 缺失 (HERV15yq1-HERV15yq2) 5 | SCOS | 罕见 |
| AZFc 臂间 缺失 (b2/b4) 5 | 少精子症 | 6% | ||
| 无精子症 | 13% | |||
| AZFb臂间& AZFb+c 缺失 (P5/proxP1, P5/distP1, P4/distP1) 5 | 无精子症 | 1%-2% | ||
| 末端AZFdeletion (常表现为假双着丝粒Y染色体并/ 重复 & 缺失 ) | 无精子症 | 罕见 | ||
| USP9Y | Direct DNA 6 | AZFa内的序列变异 | 精子生成不足 | 罕见 7 |
| DDX3Y | Direct DNA 6 | AZFa内的序列变异 8 | 若DDX3Y & USP9Y 同时缺失,为无精子症 | 罕见 |
SCOS = 唯支持细胞综合征
ND =无数据
1.
2.
3.
男性不育遗传机制引起的缺失或致病变异的发生率。
4.
编码区和侧翼内含子区的基因组DNA 序列分析不容易检测到缺失/重复;可用的检测方法有:包含该 基因/染色体片段的定量PCR,长片段PCR、多重连接探针依赖的扩增(MLPA)和 染色体微阵列(CMA) 。
5.
侧翼重复 [Kamp et al 2000, Sun et al 2000, Kamp et al 2001, Yen 2001, Oates et al 2002, Repping et al 2002]
6.
“Direct DNA” 是GeneReviews中的一个与研究测试所用的方法有关的名词,可包括 致病变异的定点分析,变异扫描, 序列分析, 缺失/ 重复检测,或其他分子遗传学检测的任意组合。在解释序列分析结果时需要考虑的问题,请点击 此处。
7.
位于AZFa区域(见 Figure 1)涉及 USP9Y基因内的致病变异已在很少的病例中有报道。 [Sun et al 1999]. 这些类型的致病变异不能通过缺失/重复分析被检测到,因此需要其他类型的检测(如, 序列分析)。
8.
检测策略
先证者诊断的确认/建立. 一个先证者诊断的确认/建立。应评估精子数量和形态学/运动性,并利用 缺失/重复分析进行分子遗传学检测。如果未发现缺失,则应使用足够覆盖Y 染色体的探针(检测拷贝数变化)进行CMA分析。如果未发现异常,应进行常规 细胞遗传学研究,以检测是否有染色体平衡重排。如果有 必要,采用G显带,Q显带和特异Y探针的 FISH等细胞遗传学分析,以明确一个复杂的重排。
对高危妊娠采用产前诊断和胚胎植入前遗传学诊断(PGD)前,需要预先明确Y 染色体长臂AZF区的缺失。
临床特点
临床描述
Y染色体不育的男性通常没有明显的症状,虽然体检可以显示小睾丸。Y染色体不育男性有无精子症或重度、中度、轻度的少精子症( Table 1 )。某些情况下少 精子症可有生育能力。
Yq靠近着丝粒区域的缺失包含一个假生长控制基因GCY ,Yq缺失的个体可能会出现身材矮小[Kirsch et al 2002, Kirsch et al 2004]。异常的身材也可由拟常染色体区域内(PAR)可影响SHOX基因的隐藏的拷贝数变化导致 [ Jorgez et al 2011]。然而,这些状况是罕见的,局限于AZF区域的跟这些异常表型无关。
基因型-表型的相关性
每个AZF区域包含几个在精子发生不同阶段发挥作用的基因。今后对不育男性这些基因的分析可得到更精确的 基因型-表型相关 性。然而,Y 染色体上大多数生育基因的多拷贝性和多态性很难精确地定义它们的作用。
当初确定的AZFb和AZFc区域,现已发现他们之间有部分重叠 (Figure 1) [Repping et al 2002]。许多文献仍然涉及这些区域;因此,作者加入了回文重复的参考信息,现在可以更精确地定义缺失[ Silber 2011 ]。
- 包含整个ZAFa区域的臂间或末端缺失是罕见的,通常导致严重的唯支持细胞综合征(SCOS)表型[ Silber 2011 ] (见 鉴别诊断)。 HERV15yq1-HERV15yq2之间重复序列的重组介导了Y染色体的臂间缺失。位于AZFa区域的一个单拷贝 基因 (USP9Y) 与不育表型直接相关,在两名精子生成不足的非SCOS不育的男性中检测到USP9Y基因的一个 单核甘酸变异和一个缺失[Sun et al 1999]。 在精子生成不足的不育患者中发现了USP9Y 的完全缺失[Luddi et al 2009],表明通常与AZFa缺失相关的SCOS不是由USP9Y基因的单独缺失导致的,而是必须包括至少一个邻近的DDX3Y基因的缺失才可以导致无精子症。涉及USP9Y和DDX3Y两个基因的AZFa完全缺失可导致比单独 USP9Y基因突变更为严重的表型。
- 包含AZFb和/或AZFb+c (以下简称为AZFb/c)区域的臂间或末端缺失是由回文重复,P5/proxP1,P5/distP1或P4/distP1,之间的 重组介导的。 这些缺失是罕见的,通常会导致严重的无精子症[Repping et al 2002, Silber 2011]。AZFb的部分缺失移除了整个P4回文序列,抑制了精母细胞的成熟,但是该缺失可以 传递[Kichine et al 2012]。
- 只包含AZFc 区域的臂间或末端缺失是由b2/b4回文重复之间的重组介导的,导致一种可变的不育 表型,从无精子症和SCOS到重度或轻度的少精子症 [Oates et al 2002, Silber 2011]。这种类型的缺失是常见的。
- b1/b3,b2/b3这两个部分的缺失,被认为是良性的拷贝数变异(多态性)[Repping et al 2003, Fernandes et al 2004, Machev et al 2004, Ferlin et al 2007]。 另一个部分的缺失, gr/gr,可对生育有影响,这取决于其种族和地理区域[ Stouffs et al 2011]。 gr/gr缺失的男性也有基因的补偿重复[ Noordam et al 2011]。
外显率
相同的Y染色体缺失偶尔引起一些男性不育,而其他男性没有表现为不育,这种情况在一个家系中是罕见的[Chang et al 1999, Saut et al 2000, Gatta et al 2002, Repping et al 2003]。这些现象被误解为是可变外显率。然而事实是,一个Y染色体AZF区域缺失的严重少精子症男性偶尔与生育能力极佳的女性怀孕所致。
患病率
Y染色体缺失和微缺失男性的患病率估计在1/2000-1/3000之间。
在无精子症或严重少精子症男性中,Yq微缺失的几率约为5% - 15% [Vogt 1997, Silber et al 1998, Oates et al 2002, Silber 2011]。
未观察到基于种族的患病率差异。然而,gr/gr缺失对生育的影响,可能会因种族和地理区域的差异而有所不同[ Stouffs et al 2011]。例如,日本男性的gr/gr缺失是非常常见的(25%),这仅是男性不育的一个“风险因素”。
遗传相关疾病
除了不育,没有其他疾病认为是由AZF区域Y染色体长臂的缺失所致。 Y染色体缺失不育男性的健康状况是良好。然而,复杂的染色体重排可导致拟常染色体区(PAR)内的基因中断,例如SHOX,出现额外的表型,如身材矮小[ Jorgez et al 2011]。
鉴别诊断
15% - 20%的育龄夫妇受不孕不育的影响。男性因素的不育约占一半,但是这个经常被引用的数字是缺乏证据的。最可能的是,严重的少精子症足以导致不孕不育的夫妇仅有10%。 除了Y染色体缺失以外,男性不育的原因很多而且经常引起争议。在大多数情况下,男性不育症病因不明。除了Y染色体缺失以外,男性不育的可能原因 包括以下几种情况:
- 射精管梗阻,体检可以评估[Practice Committee of the American Society for Reproductive Medicine 2004]。在本评估中应考虑先天性双侧输精管缺如(见 CFTR-相关疾病)。 CFTR相关疾病包括囊性纤维化(CF)和先天性双侧输精管缺如(CBAVD)。所有CF男性的不育是由Wolffian管的缺失、萎缩、纤维化引起的少精子症导致。CBAVD 多见于无肺和胃肠症状的CF男性,他们有一个严重的CFTR致病突变等位基因和另一个轻度的T5突变等位基因。 这些人的CTFR致病突变仍可产生正常量10%的蛋白,足以防止临床CF,但是不够胎儿Wolffian管发育的剂量[ Chillón et al 1995]。无精子症男性是不育的。CF是常染色体隐性遗传的。
- 抗精子抗体引起的免疫异常(有争议)
- 感染(如,流行性腮腺炎性睾丸炎、附睾炎、尿道炎); 一般可通过既往史与Y 染色体不育相鉴别。
- 血管异常(精索静脉曲张);可通过体检确诊,但其与男性不育的相关性已受到大多数生殖内分泌学家的强烈质疑,是非常有争议的[ Silber 2001 ]。
- 创伤(通过病史可鉴别)
- 内分泌异常;也很罕见(例如,先天性肾上腺皮质增生症、 孤立的卵泡刺激素(FSH)不足,和高泌乳素血症)。这些可通过激素检测来鉴别。 卡尔曼综合征(KS),与GnRH缺陷(IGD)和嗅觉缺失相关,需要考虑。一些KS男性新生儿时期有小阴茎和隐睾。KS成人有第二性征的不全发育和青春期睾丸体积(<4ml)。 KAL1与FGFR1是已知仅有的与卡尔曼综合征相关的两个基因。在这两个基因中,致病突变约占 KS的15% - 25%。非生殖表型:
- KAL1致病变异的男性中。手指足趾的联带运动(镜像运动),单侧肾缺如,感音神经性听力缺失,硬腭高拱
- FGFR1致病变异的男性中。 手指足趾的联带运动,唇腭裂,牙齿发育不全,短、并指畸形,胼胝体发育不全
- 睾丸肿瘤,或因接触有毒物质而引起的其他肿瘤
- 暴露于有毒物质 如放射、化疗、热暴露(完整的病史评估)
- Klinefelter综合征 (XXY),细胞遗传学分析或CMA可检测,在非梗阻性无精子症和少精子症男性中约占4%。
- 平衡的染色体重排, 约1.5%的非梗阻性无精子症和少精子症男性,可通过 细胞遗传学评估检测。在这种情况下,也可能有多次流产和/或各种表型异常的家族史。
唯支持细胞综合征 (SCOS) 是用于描述男性生殖发育不全的一个术语。有很多致病原因,包括Y 缺失,暴露于有毒的化疗药物或辐射,腮腺炎性睾丸炎, 唐氏综合征,克氏综合征(47,XXY), 先天性肾上腺发育不全,孤立的卵泡刺激素(FSH)不足和高泌乳素血症。对于每一种病因,应通过病史、其他异常或 症状的存在,或 染色体分析与Y染色体不育鉴别。在大多数情况下,SCOS的病因是未知的。
管理
初步诊断后的评估
为了确定一个诊断为Y染色体不育症的患者的疾病程度,建议进行精液分析,以确定精子的数量、活力和形态。
对症治疗
男性Y染色体不育的夫妇可以选择ICSI(卵胞浆内单精子注射)进行体外受精[ Silber 2011 ]。在这个过程中,精子从射精(男性少精症)或睾丸活检(男性无精子症)获取,再通过IVF(体外受精)注入收获的卵子中。
大多数AZFc缺失的男性可成功获精,但是AZFb或AZFa缺失的男性鲜有成功。这是因为 常染色体DAZ拷贝(DAZL)可以作为一个“备份基因”,有助于因AZFc缺失而使 DAZ基因移除的男性有少量残留的的精子生成。AZFa和AZFb没有这样的常染色体的“备份”存在。
文献中一直对SCOS(唯支持细胞综合征)的定义有困惑。非梗阻性无精子症(NOA)有两个主要原因:发育停滞和唯支持细胞。随着发育停滞,精母细胞 不能完成减数分裂I。但是在60%的病例中,少数精母细胞可进一步发育成精子,可以从睾丸获取。同样,约60%SCOS病例的少数小管中包含从小的精子生成 灶产生的少许的精子。
在ICSI和IVF受精之前,讨论Y染色体不育在男性后代中传递的可能性是很重要的(见 遗传咨询)[Stouffs et al 2005]。
在可获精的男性中,Y染色体长臂缺失的存在对受精或妊娠率没有明显的影响[ Oates et al 2002, Kihaile et al 2004, Silber 2011]。 出生缺陷的风险与其他通过辅助生殖技术怀孕的不孕夫妇没有什么不同[ Choi et al 2004, Davies et al 2012]。
研究中的治疗
搜索 ClinicalTrials.gov 获取各种疾病的临床研究信息。注意:这种疾病可能没有临床试验。
其他
睾丸取精体外受精对于AZFb和AZFa缺失的男性是无效,但对大多数AZFc缺失男性可获得成功[ Oates et al 2002, Stouffs et al 2005, Reyes-Vallejo et al 2006, Silber 2011]。
遗传咨询
遗传咨询是向个人和家庭提供关于遗传病的性质,传递和影响信息的过程,以帮助他们做出明智的医疗和个人决定。 以下部分涉及遗传风险评估和家族史和遗传检测的应用,以明确家庭成员的遗传状况。 本节不能涉及个人所面临的所有的个人,文化或伦理问题,也不能替代遗传学专业人士的咨询。 —ED.
遗传方式
Y染色体不育以Y-连锁方式遗传。
家庭成员的风险
父母/先证者的父亲
- Y染色体AZFc缺失的男性生育一个不育儿子的家庭是罕见的[ Chang et al 1999, Saut et al 2000, Gatta et al 2002, Repping et al 2003]。然而,应该强调的是,在这些家庭中,AZFc缺失的父亲都是严重少精子的,虽然他的精子数很低,但是在有一个非常年轻且生育能力极佳的伴侣的情况下,还是可以孕育的。
- gr/gr 缺失的男性有可变的生育力,可以很容易地将缺失传递给下一代[ Silber 2011 ]。b1/b3和b2/b3 缺失是无足轻重的,对男性生育没有影响。
同胞/先证者的兄弟
- 在非常罕见的情况下,当家庭内有AZFc区相同缺失可能导致一些人不育而其他人正常,这种情况下, 先证者的兄弟可能会有不育的风险[Chang et al 1999, Saut et al 2000, Repping et al 2003]。然而,需要再次澄清的是,AZFc缺失的男性都有严重的精子生成缺陷, 仅当其伴侣有极佳的生育能力时才可补偿其生育缺陷。gr/gr缺失个体的生育率是可变的。
先证者的后代
- 男性胎儿有与其父相同的Y染色体缺失时,有很高的不育风险[ Page et al 1999, Oates et al 2002, Silber 2011]。目前尚无Y染色体缺失的生殖细胞嵌合的报道;因此,Y缺失总是传递给男性后代。
其他家庭成员. 由于Y染色体缺失是正常精子生成能力父亲睾丸中1/2000个精子发生的 新生事件,所以大家族成员不大可能发生Y染色体不育。此外,对AZF缺失传递的可能性是极小的。
相关的遗传咨询问题
生育计划
- 孕前是确定遗传风险和讨论产前诊断可行性的最佳时机。
DNA存储是对将来可能用到的DNA(通常从白细胞提取)进行存储。因为将来我们可能对检测方式和对基因、等位基因变异和疾病的理解有所改进,应考虑存储患者的DNA。
产前检测
Y染色体缺失男性通过辅助生殖技术(ART)妊娠的产前诊断,是可以通过羊膜穿刺术取胎儿细胞(通常在~ 孕15-18周进行)或绒毛膜绒毛取样(通常在~ 孕10-12周进行)的DNA分析来进行的。检测包括确定胎儿的性别和/或Y染色体缺失是否存在。对于由父源Y染色体重排(如,假双着丝粒Y或环型Y)导致的染色体异常(如,45,X 嵌合)的高危妊娠,产前诊断也是可行的。
注:孕龄以月经期计算为准,从末次月经的第一天开始,也可通过超声测量。
资源
GeneReviews的工作人员选了以下疾病特异的和/或保护支持组织和/或为患此疾病的个人及其家人的利益提供登记。 GeneReviews不对其他组织提供的信息负责。有关选择标准的信息,请点击此处。
- InterNational Council on Infertility Information Dissemination, Inc. (INCIID)5765 F Burke Centre PkwyBox 330Burke VA 22015Email: inciidinfo@inciid.org
- RESOLVE: The National Infertility Association7918 Jones Branch DriveSuite 300McLean VA 22102Phone: 703-556-7172Fax: 703-506-3266Email: info@resolve.org
分子遗传学
分子遗传学和OMIM表格中的信息可能与GeneReview中的其他信息有不同:表中可能包含了更能多最近的信息 —ED.
Table A.
Y染色体不育:基因和数据库
Table B.
OMIM Entries for Y Chromosome Infertility (View All in OMIM)
| 400003 | DELETED IN AZOOSPERMIA; DAZ |
| 400005 | UBIQUITIN-SPECIFIC PROTEASE 9, Y CHROMOSOME; USP9Y |
| 400006 | RNA-BINDING MOTIF PROTEIN, Y CHROMOSOME, FAMILY 1, MEMBER A1; RBMY1A1 |
| 400010 | DEAD/H BOX 3, Y-LINKED; DDX3Y |
| 400012 | VARIABLY CHARGED, Y CHROMOSOME; VCY |
| 400013 | BASIC PROTEIN, Y CHROMOSOME, 2; BPY2 |
| 400015 | XK-RELATED PROTEIN ON Y CHROMOSOME |
| 400016 | CHROMODOMAIN PROTEIN, Y CHROMOSOME, 1; CDY1 |
| 400018 | CHROMODOMAIN PROTEIN, Y CHROMOSOME, 2; CDY2 |
| 400019 | PTPBL-RELATED GENE ON Y; PRY |
| 400026 | DELETED IN AZOOSPERMIA 2; DAZ2 |
| 400027 | DELETED IN AZOOSPERMIA 3; DAZ3 |
| 400029 | HEAT-SHOCK TRANSCRIPTION FACTOR, Y-LINKED; HSFY |
| 400030 | RIBOSOMAL PROTEIN S4, Y-LINKED, 2; RPS4Y2 |
| 400041 | PTPBL-RELATED GENE ON Y, 2; PRY2 |
| 400042 | SPERMATOGENIC FAILURE, Y-LINKED, 1; SPGFY1 |
| 415000 | SPERMATOGENIC FAILURE, Y-LINKED, 2; SPGFY2 |
| 426000 | LYSINE-SPECIFIC DEMETHYLASE 5D; KDM5D |
分子及遗传学发病机制
座位名称. 最初,无精子症的三个区域(a, b, and c)定义为AZFa, AZFb和AZFc。然而,由于AZFb和AZFc存在重叠,就用了一个新的术语根据侧翼回文重复名称来区分缺失[ Repping et al 2002, Silber 2011]。很可能会有其他的缺失类型被发现(见Table 2)。
基因. 参照NCBI列出了定位于Y染色体长臂的基因,基因功能注释保持当前。
染色体座位. 男性不育表型映射到Y 染色体上相对广泛的区域 ( Figure 1). 这些区域是根据不育男性的 缺失间隔定义的。 这些大量的臂间缺失是由Y染色体回文重复之间的 重组介导的。
AZFa区域长度是792 kb,在Y染色体着丝粒的远端(Figure 1) [Kamp et al 2001]。 位于AZFa附近的两个 HERV15 (HERV15yq1-HERV15yq2) 病毒序列已被确定为AZFa缺失发生的机制[Kamp et al 2000, Sun et al 2000, Kamp et al 2001]。
AZFb和AZFc区域之间存在重叠,远侧的AZFc位于Y染色体异染色质带Yq12 的近端(Figure 1)。最常见的Y染色体 缺失,原来称为AZFc缺失,移除了回文序列b2和b4之间3.5M的区域[Kuroda-Kawaguchi et al 2001, Vogt 2005]。AZFb/c区域两个最大的臂间缺失移除了回文序列P5和近端P1之间6.2M的区域(原来称为AZFb缺失),或者移除了回文序列P5和远端P1之间7.7M的区域(原来称为AZFa和AZFc缺失)[ Repping et al 2002]。另一个不太常见的缺失移除了回文序列P4和远端P1之间7M的区域。 涉及回文序列gr/gr,b1/b3或b2/b3的AZFc中的小缺失,通常被认为是良性的,但是可能在某些民族中与不育有相关性( Figure 1) [Stouffs et al 2011]。 AZFb中的部分缺失移除了整个P4回文序列,明显降低了精母细胞的成熟,是可以传递[ Kichine et al 2012]。
通过细胞遗传学和PCR分析,Y缺失子集的检测似乎已近终点。这些缺失往往与严重的精子发生受损相关;根据断裂点分析,这些缺失全部或部分移除了Y 染色体的末端q12带的AZF区域。这些所谓的“末端缺失”通常不是末端,而是表现为更复杂的重排,如等臂双着丝粒Y染色体,它通常是不稳定且与45,X嵌合细胞相关。事实上,Turner综合征(45,X)可由一个等臂双着丝粒Y染色体缺失导致[ Lange et al 2009]。
Y染色体缺失可与拟常染色体区域(PAR)的缺陷有关,导致PAR区域基因的拷贝数变化,如 SHOX基因,是一个重要的身高基因[Jorgez et al 2011]。这些异常在Y染色体的复杂重排中很普遍,可以通过CMA分析来检测。
到目前为止,通过两个个体中基因有限的致病变异的研究,发现只有一个单拷贝 基因,USP9Y,与不育表型相关[ Sun et al 1999]。然而,此基因的完全缺失与轻度的精子生成减少和生育相关[ Luddi et al 2009]。应当注意的是,许多Y连锁基因是多拷贝的,因此致病变异的解释就变得有困难。
该区域内的感兴趣基因. 正常的Y染色体包含所有的AZF区域。Y染色体完整的 序列分析和Y连锁基因的列表已被报道[Skaletsky et al 2003, Silber 2011, Hughes & Rozen 2012, Rozen et al 2012]。Y染色体AZF区域的大多数基因都是在睾丸组织中特异表达的,是男性生育功能候选基因。其他未知的转录本也能在男性生育中发挥作用。 USP9Y是唯一的单 拷贝 基因,在一位不育的男性中发现该基因的 致病变异,显示USP9Y基因与男性生育有直接的关系[Sun et al 1999]。
读者可从NCBI网站查询当前Y连锁基因的注释。下面是一些被认为在男性生育中起作用的基因的简短描述。从 基因的定位,在生殖细胞中的表达和/或其他物种生育相关基因的同源性来看,这些基因在疾病中的作用仍然 是不确定的。应该指出的是,Y 染色体上的几个Y连锁基因是多拷贝的;多个拷贝的存在使他们在男性生育中的作用更加复杂。
- USP9Y 是一个单拷贝基因,定位于近端AZFa区。 USP9Y与其X连锁的同源基因USP9X编码与泛素特异性蛋白酶类似的蛋白。两位不育男性中的 USP9Y变异,提示该基因是男性生育直接牵涉基因[Sun et al 1999]。然而,该基因的完全缺失致轻度的生精障碍且有生育能力[ Luddi et al 2009]。
- DDX3Y是定位于AZFa区域的一个单拷贝基因,X染色体上有其同源基因,DDX3。它们编码广泛表达的DEAD盒蛋白,但是 DDX3Y在睾丸组织中表达一个可变转录本[Lahn & Page 1997]。仅在雄性生殖细胞,主要是精原细胞中,可以检测到 DDX3Y的翻译[Ditton et al 2004]。
- VCY是一个多拷贝基因家族,在X染色体上有同源基因 (VCX, VCX2)。VCY (BPY1)和BPY2 (VCY2) ,定位于AZFb/c区域,在雄性生殖细胞中表达, BPY2在精原细胞、精母细胞和圆形精子细胞中特异表达[Lahn & Page 2000, Tse et al 2003]。
- 定位于AZFb区域的HSFY1和HSFY2,及其位于X染色体上的同源基因HSFX1是热休克转录因子家族的成员。HSFY1 和 HSFY2 仅在支持细胞和生精细胞中表达[Shinka et al 2004, Vinci et al 2005]。 HSFY1和HSFY2 的缺失可致精母细胞成熟略有减少[Kichine et al 2012]。
- 位于AZFb区域的RPS4Y2基因编码一个睾丸特异表达的核糖体蛋白。相反,Y连锁的RPS4Y1基因(位于Y染色体短臂)和 X连锁的同源基因RPS4X,是广泛表达的[Lopes et al 2010]。
- RBMY是一个定位于Y染色体短臂、长臂的AZFb及其他区域的多拷贝 基因家族。大约有15-20个RBMY的拷贝,但仅有6个是有功能的。有一个X连锁的同源基因,RBMX,是广泛表达的[ Delbridge et al 1999]。 RBMY家族的基因仅在睾丸中表达,生成可与RNA结合的蛋白,调控 剪接因子的活性[Skrisovska et al 2007, Dreumont et al 2010]。
- PRY是一个多拷贝基因家族,有四个成员。PRY1/2 定位于AZFb , PRY3/4定位于AZFc。这些基因编码的蛋白与蛋白酪氨酸磷酸酶有低度的相似性,非受体型13,可能参与受损的精子和精子细胞的凋亡 [Stouffs et al 2004]。
- DAZ是一个多拷贝基因家族,定位于AZFc区域。DAZ基因家族由包含4个拷贝(DAZ1-4)的重复基因簇组成[ Saxena et al 2000]。在人类3号染色体上有一个DAZ的同源基因( DAZL)。它们编码在精原细胞中表达的RNA结合蛋白,并参与单倍体配子的发育调控[ Menke et al 1997, Kee et al 2009]。
- CDY是一个多拷贝基因家族,包含四个有显著功能的基因拷贝(CDY1, CDY1B, CDY2A和CDY2B),定位于AZFb/c。CDY家族基因代表来自 常染色体基因(CDYL) 的加工处理的转座子,在猿系Y染色体上是扩大的。CDY家族基因编码的染色质蛋白在成熟的精子细胞中特异表达,可以促进组蛋白向 鱼精蛋白的过渡[ Lahn et al 2002, Dorus et al 2003]。
缺失. 与男性不育相关的Y染色体异常导致基因的 缺失。Y染色体缺失范围通常很大,从而导致多个基因的缺失。Y染色体长臂的几个区域受到牵连( Figure 1)。
注:除单基因条目,OMIM列出了两个Y缺失综合征:
- 415000 生精障碍,Y连锁的,2 (SPGFY2),指AZFa,b或c的缺失
见 Table B.
正常基因产物. 不育男性AZF缺失区域中包含了几个基因。然而,正如上面所讨论的,它们在正常精子发生中的作用尚不完全清楚。当前Y连锁基因列表及其蛋白产物的术语注释请参照 NCBI。
异常基因产物. Y染色体缺失导致基因产物的缺失。这些缺失通常包括一个以上的基因或多拷贝的基因。就 USP9Y基因来说,局限于基因内的4bp缺失被发现可导致一个 外显子跳跃和截短蛋白的生成[Sun et al 1999]。
参考文献
发布的指南/政策声明
- Practice Committee of the American Society for Reproductive Medicine. Report on evaluation of the azoospermic male. Fertil Steril. 2004;82 Suppl 1:S131–6. [PubMed: 15363709]
引用文献
- Akimoto C, Kitagawa H, Matsumoto T, Kato S. Spermatogenesis-specific association of SMCY and MSH5. Genes Cells. 2008;13:623–33. [PubMed: 18459961]
- Aknin-Seifer IE, Touraine RL, Faure AK, Fellmann F, Chouteau J, Levy R. Two fast methods for detection of Y-microdeletions. Fertil Steril. 2005;84:740–2. [PubMed: 16169411]
- Bosch E, Jobling MA. Duplications of the AZFa region of the human Y chromosome are mediated by homologous recombination between HERVs and are compatible with male fertility. Hum Mol Genet. 2003;12:341–7. [PubMed: 12554687]
- Chang PL, Sauer MV, Brown S. Y chromosome microdeletion in a father and his four infertile sons. Hum Reprod. 1999;14:2689–94. [PubMed: 10548602]
- Chillón M, Casals T, Mercier B, Bassas L, Lissens W, Silber S, Romey MC, Ruiz-Romero J, Verlingue C, Claustres M, Nunes V, Férec C, Estivill X. Mutations in the cystic fibrosis gene in patients with congenital absence of the vas deferens. N Engl J Med. 1995;332:1475–80. [PubMed: 7739684]
- Choi JM, Chung P, Veeck L, Mielnik A, Palermo GD, Schlegel PN. AZF microdeletions of the Y chromosome and in vitro fertilization outcome. Fertil Steril. 2004;81:337–41. [PubMed: 14967370]
- Davies MJ, Moore VM, Willson KJ, Van Essen P, Priest K, Scott H, Haan EA, Chan A. Reproductive technologies and the risk of birth defects. N Engl J Med. 2012;366:1803–13. [PubMed: 22559061]
- Delbridge ML, Lingenfelter PA, Disteche CM, Graves JA. The candidate spermatogenesis gene RBMY has a homologue on the human X chromosome. Nat Genet. 1999;22:223–4. [PubMed: 10391206]
- Ditton HJ, Zimmer J, Kamp C, Rajpert-De Meyts E, Vogt PH. The AZFa gene DBY (DDX3Y) is widely transcribed but the protein is limited to the male germ cells by translation control. Hum Mol Genet. 2004;13:2333–41. [PubMed: 15294876]
- Dorus S, Gilbert SL, Forster ML, Barndt RJ, Lahn BT. The CDY-related gene family: coordinated evolution in copy number, expression profile and protein sequence. Hum Mol Genet. 2003;12:1643–50. [PubMed: 12837688]
- Dreumont N, Bourgeois CF, Lejeune F, Liu Y, Ehrmann IE, Elliott DJ, Stevenin J. Human RBMY regulates germline-specific splicing events by modulating the function of the serine/arginine-rich proteins 9G8 and Tra2-beta. J Cell Sci. 2010;123:40–50. [PubMed: 20016065]
- Ferlin A, Arredi B, Speltra E, Cazzadore C, Selice R, Garolla A, Lenzi A, Foresta C. Molecular and clinical characterization of y chromosome microdeletions in infertile men: a 10-year experience in Italy. J Clin Endocrinol Metab. 2007;92:762–70. [PubMed: 17213277]
- Fernandes S, Paracchini S, Meyer LH, Floridia G, Tyler-Smith C, Vogt PH. A large AZFc deletion removes DAZ3/DAZ4 and nearby genes from men in Y haplogroup N. Am J Hum Genet. 2004;74:180–7. [PMC free article: PMC1181906] [PubMed: 14639527]
- Gatta V, Stuppia L, Calabrese G, Morizio E, Guanciali-Franchi P, Palka G. A new case of Yq microdeletion transmitted from a normal father to two infertile sons. J Med Genet. 2002;39:E27. [PMC free article: PMC1735151] [PubMed: 12070259]
- Giachini C, Laface I, Guarducci E, Balercia G, Forti G, Krausz C. Partial AZFc deletions and duplications: clinical correlates in the Italian population. Hum Genet. 2008;124:399–410. [PubMed: 18807255]
- Hughes JF, Rozen S. Genomics and genetics of human and primate y chromosomes. Annu Rev Genomics Hum Genet. 2012;13:83–108. [PubMed: 22483277]
- Jorgez CJ, Weedin JW, Sahin A, Tannour-Louet M, Han S, Bournat JC, Mielnik A, Cheung SW, Nangia AK, Schlegel PN, Lipshultz LI, Lamb DJ. Aberrations in pseudoautosomal regions (PARs) found in infertile men with Y-chromosome microdeletions. J Clin Endocrinol Metab. 2011;96:E674–9. [PMC free article: PMC3070254] [PubMed: 21252244]
- Kamp C, Hirschmann P, Voss H, Huellen K, Vogt PH. Two long homologous retroviral sequence blocks in proximal Yq11 cause AZFa microdeletions as a result of intrachromosomal recombination events. Hum Mol Genet. 2000; 9:2563–72. [PubMed: 11030762]
- Kamp C, Huellen K, Fernandes S, Sousa M, Schlegel PN, Mielnik A, Kleiman S, Yavetz H, Krause W, Küpker W, Johannisson R, Schulze W, Weidner W, Barros A, Vogt PH. High deletion frequency of the complete AZFa sequence in men with Sertoli-cell-only syndrome. Mol Hum Reprod. 2001;7:987–94. [PubMed: 11574668]
- Kee K, Angeles VT, Flores M, Nguyen HN, Reijo Pera RA. Human DAZL, DAZ and BOULE genes modulate primordial germ-cell and haploid gamete formation. Nature. 2009;462:222–5. [PMC free article: PMC3133736] [PubMed: 19865085]
- Kichine E, Rozé V, Di Cristofaro J, Taulier D, Navarro A, Streichemberger E, Decarpentrie F, Metzler-Guillemain C, Lévy N, Chiaroni J, Paquis-Flucklinger V, Fellmann F, Mitchell MJ. HSFY genes and the P4 palindrome in the AZFb interval of the human Y chromosome are not required for spermatocyte maturation. Hum Reprod. 2012;27:615–24. [PubMed: 22158087]
- Kihaile PE, Kisanga RE, Aoki K, Kumasako Y, Misumi J, Utsunomiya T. Embryo outcome in Y-chromosome microdeleted infertile males after ICSI. Mol Reprod Dev. 2004;68:176–81. [PubMed: 15095338]
- Kirsch S, Weiss B, Schön K, Rappold GA. The definition of the Y chromosome growth-control gene (GCY) critical region: relevance of terminal and interstitial deletions. J Pediatr Endocrinol Metab. 2002; 15 Suppl 5:1295–300. [PubMed: 12510983]
- Kirsch S, Weiss B, Zumbach K, Rappold G. Molecular and evolutionary analysis of the growth-controlling region on the human Y chromosome. Hum Genet. 2004;114:173–81. [PubMed: 14579146]
- Kuroda-Kawaguchi T, Skaletsky H, Brown LG, Minx PJ, Cordum HS, Waterston RH, Wilson RK, Silber S, Oates R, Rozen S, Page DC. The AZFc region of the Y chromosome features massive palindromes and uniform recurrent deletions in infertile men. Nat Genet. 2001;29:279–86. [PubMed: 11687796]
- Lahn BT, Page DC. Functional coherence of the human Y chromosome. Science. 1997;278:675–80. [PubMed: 9381176]
- Lahn BT, Page DC. A human sex-chromosomal gene family expressed in male germ cells and encoding variably charged proteins. Hum Mol Genet. 2000;9:311–9. [PubMed: 10607842]
- Lahn BT, Tang ZL, Zhou J, Barndt RJ, Parvinen M, Allis CD, Page DC. Previously uncharacterized histone acetyltransferases implicated in mammalian spermatogenesis. Proc Natl Acad Sci U S A. 2002;99:8707–12. [PMC free article: PMC124363] [PubMed: 12072557]
- Lange J, Skaletsky H, van Daalen SK, Embry SL, Korver CM, Brown LG, Oates RD, Silber S, Repping S, Page DC. Isodicentric Y chromosomes and sex disorders as byproducts of homologous recombination that maintains palindromes. Cell. 2009; 138:855–69. [PMC free article: PMC3616640] [PubMed: 19737515]
- Lopes AM, Miguel RN, Sargent CA, Ellis PJ, Amorim A, Affara NA. The human RPS4 paralogue on Yq11.223 encodes a structurally conserved ribosomal protein and is preferentially expressed during spermatogenesis. BMC Mol Biol. 2010;11:33. [PMC free article: PMC2884166] [PubMed: 20459660]
- Luddi A, Margollicci M, Gambera L, Serafini F, Cioni M, De Leo V, Balestri P, Piomboni P. Spermatogenesis in a man with complete deletion of USP9Y. N Engl J Med. 2009;360:881–5. [PubMed: 19246359]
- Machev N, Saut N, Longepied G, Terriou P, Navarro A, Levy N, Guichaoua M, Metzler-Guillemain C, Collignon P, Frances AM, Belougne J, Clemente E, Chiaroni J, Chevillard C, Durand C, Ducourneau A, Pech N, McElreavey K, Mattei MG, Mitchell MJ. Sequence family variant loss from the AZFc interval of the human Y chromosome, but not gene copy loss, is strongly associated with male infertility. J Med Genet. 2004;41:814–25. [PMC free article: PMC1735624] [PubMed: 15520406]
- Menke DB, Mutter GL, Page DC. Expression of DAZ, an azoospermia factor candidate, in human spermatogonia. Am J Hum Genet. 1997;60:237–41. [PMC free article: PMC1712565] [PubMed: 8981970]
- Noordam MJ, Westerveld GH, Hovingh SE, van Daalen SK, Korver CM, van der Veen F, van Pelt AM, Repping S. Gene copy number reduction in the azoospermia factor c (AZFc) region and its effect on total motile sperm count. Hum Mol Genet. 2011;20:2457–63. [PubMed: 21429917]
- Oates RD, Silber S, Brown LG, Page DC. Clinical characterization of 42 oligospermic or azoospermic men with microdeletion of the AZFc region of the Y chromosome, and of 18 children conceived via ICSI. Hum Reprod. 2002;17:2813–24. [PubMed: 12407032]
- Page DC, Silber S, Brown LG. Men with infertility caused by AZFc deletion can produce sons by intracytoplasmic sperm injection, but are likely to transmit the deletion and infertility. Hum Reprod. 1999; 14:1722–6. [PubMed: 10402375]
- Practice Committee of the American Society for Reproductive Medicine. Report on evaluation of the azoospermic male. Fertil Steril. 2004;82 Suppl 1:S131–6. [PubMed: 15363709]
- Repping S, Skaletsky H, Brown L, van Daalen SK, Korver CM, Pyntikova T, Kuroda-Kawaguchi T, de Vries JW, Oates RD, Silber S, van der Veen F, Page DC, Rozen S. Polymorphism for a 1.6-Mb deletion of the human Y chromosome persists through balance between recurrent mutation and haploid selection. Nat Genet. 2003;35:247–51. [PubMed: 14528305]
- Repping S, Skaletsky H, Lange J, Silber S, Van Der Veen F, Oates RD, Page DC, Rozen S. Recombination between palindromes P5 and P1 on the human Y chromosome causes massive deletions and spermatogenic failure. Am J Hum Genet. 2002;71:906–22. [PMC free article: PMC419997] [PubMed: 12297986]
- Reyes-Vallejo L, Lazarou S, Morgentaler A. Y chromosome microdeletions and male infertility: who should be tested and why? BJU Int. 2006;97:441–3. [PubMed: 16469004]
- Rozen SG, Marszalek JD, Irenze K, Skaletsky H, Brown LG, Oates RD, Silber SJ, Ardlie K, Page DC. AZFc deletions and spermatogenic failure: a population-based survey of 20,000 chromosomes. Am J Hum Genet. 2012; 91:890–6. [PMC free article: PMC3487143] [PubMed: 23103232]
- Saut N, Terriou P, Navarro A, Levy N, Mitchell MJ. The human Y chromosome genes BPY2, CDY1 and DAZ are not essential for sustained fertility. Mol Hum Reprod. 2000;6:789–93. [PubMed: 10956550]
- Saxena R, de Vries JW, Repping S, Alagappan RK, Skaletsky H, Brown LG, Ma P, Chen E, Hoovers JM, Page DC. Four DAZ genes in two clusters found in the AZFc region of the human Y chromosome. Genomics. 2000; 67:256–67. [PubMed: 10936047]
- Shinka T, Sato Y, Chen G, Naroda T, Kinoshita K, Unemi Y, Tsuji K, Toida K, Iwamoto T, Nakahori Y. Molecular characterization of heat shock-like factor encoded on the human Y chromosome, and implications for male infertility. Biol Reprod. 2004;71:297–306. [PubMed: 15044259]
- Silber SJ. The varicocele dilemma. Hum Reprod Update. 2001;7:70–7. [PubMed: 11212078]
- Silber SJ. The Y chromosome in the era of intracytoplasmic sperm injection: a personal review. Fertil Steril. 2011;95:2439–48. [PubMed: 21704208]
- Silber SJ, Alagappan R, Brown LG, Page DC. Y chromosome deletions in azoospermic and severely oligozoospermic men undergoing intracytoplasmic sperm injection after testicular sperm extraction. Hum Reprod. 1998;13:3332–7. [PubMed: 9886509]
- Simoni M, Bakker E, Krausz C. EAA/EMQN best practice guidelines for molecular diagnosis of y-chromosomal microdeletions. State of the art 2004. Int J Androl. 2004;27:240–9. [PubMed: 15271204]
- Skaletsky H, Kuroda-Kawaguchi T, Minx PJ, Cordum HS, Hillier L, Brown LG, Repping S, Pyntikova T, Ali J, Bieri T, Chinwalla A, Delehaunty A, Delehaunty K, Du H, Fewell G, Fulton L, Fulton R, Graves T, Hou SF, Latrielle P, Leonard S, Mardis E, Maupin R, McPherson J, Miner T, Nash W, Nguyen C, Ozersky P, Pepin K, Rock S, Rohlfing T, Scott K, Schultz B, Strong C, Tin-Wollam A, Yang SP, Waterston RH, Wilson RK, Rozen S, Page DC. The male-specific region of the human Y chromosome is a mosaic of discrete sequence classes. Nature. 2003;423:825–37. [PubMed: 12815422]
- Skrisovska L, Bourgeois CF, Stefl R, Grellscheid SN, Kister L, Wenter P, Elliott DJ, Stevenin J, Allain FH. The testis-specific human protein RBMY recognizes RNA through a novel mode of interaction. EMBO Rep. 2007;8:372–9. [PMC free article: PMC1852761] [PubMed: 17318228]
- Stouffs K, Lissens W, Tournaye H, Haentjens P. What about gr/gr deletions and male infertility? Systematic review and meta-analysis. Hum Reprod Update. 2011;17:197–209. [PubMed: 20959348]
- Stouffs K, Lissens W, Tournaye H, Van Steirteghem A, Liebaers I. The choice and outcome of the fertility treatment of 38 couples in whom the male partner has a Yq microdeletion. Hum Reprod. 2005; 20:1887–96. [PubMed: 15790609]
- Stouffs K, Lissens W, Verheyen G, Van Landuyt L, Goossens A, Tournaye H, Van Steirteghem A, Liebaers I. Expression pattern of the Y-linked PRY gene suggests a function in apoptosis but not in spermatogenesis. Mol Hum Reprod. 2004;10:15–21. [PubMed: 14665702]
- Sun C, Skaletsky H, Birren B, Devon K, Tang Z, Silber S, Oates R, Page DC. An azoospermic man with a de novo point mutation in the Y-chromosomal gene USP9Y. Nat Genet. 1999;23:429–32. [PubMed: 10581029]
- Sun C, Skaletsky H, Rozen S, Gromoll J, Nieschlag E, Oates R, Page DC. Deletion of azoospermia factor a (AZFa) region of human Y chromosome caused by recombination between HERV15 proviruses. Hum Mol Genet. 2000; 9:2291–6. [PubMed: 11001932]
- Tse JY, Wong EY, Cheung AN, Tam WSOPC, Yeung WS. Specific expression of VCY2 in human male germ cells and its involvement in the pathogenesis of male infertility. Biol Reprod. 2003;69:746–51. [PubMed: 12724276]
- Vinci G, Raicu F, Popa L, Popa O, Cocos R, McElreavey K. A deletion of a novel heat shock gene on the Y chromosome associated with azoospermia. Mol Hum Reprod. 2005;11:295–8. [PubMed: 15734897]
- Vogt PH. Genetics of idiopathic male infertility: Y chromosomal azoospermia factors (AZFa, AZFb, AZFc). Baillieres Clin Obstet Gynaecol. 1997;11:773–95. [PubMed: 9692017]
- Vogt PH. Azoospermia factor (AZF) in Yq11: towards a molecular understanding of its function for human male fertility and spermatogenesis. Reprod Biomed Online. 2005;10:81–93. [PubMed: 15705299]
- WHO. WHO Laboratory Manual for the Examination and Processing of Human Semen. 5 ed. Geneva, Switzerland: World Health Organization; 2010. Available online. Accessed 8-24-16.
- Yen P. The fragility of fertility. Nat Genet. 2001;29:243–4. [PubMed: 11687786]
推荐阅读
- Jungwirth A, Giwercman A, Tournaye H, Diemer T, Kopa Z, Dohle G, Krausz C., EAU Working Group on Male Infertility. European Association of Urology guidelines on Male Infertility: the 2012 update. Eur Urol. 2012; 62:324–32. [PubMed: 22591628]
- Krausz C. Male infertility: pathogenesis and clinical diagnosis. Best Pract Res Clin Endocrinol Metab. 2011;25:271–85. [PubMed: 21397198]
- Massart A, Lissens W, Tournaye H, Stouffs K. Genetic causes of spermatogenic failure. Asian J Androl. 2012;14:40–8. [PMC free article: PMC3735159] [PubMed: 22138898]
章注
修订历史
- 18 October 2012 (me) Comprehensive update posted live
- 19 March 2007 (me) Comprehensive update posted to live Web site
- 28 February 2006 (cmd) Revision: Diagnosis: Cytogenetic analysis
- 16 September 2004 (me) Comprehensive update posted to live Web site
- 6 February 2004 (cd) Revision: change in 基因 name
- 31 October 2002 (me) Review posted to live Web site
- January 2001 (cmd) Original submission